Propagation – a bottleneck in tree breeding programmes?
Paul Schroeder and Clemens Altaner, New Zealand Tree Grower November 2016.
Large gains can be expected from tree breeding. You just need to compare wild corn, banana or wheat with what is grown commercially today. An undesirable feature of tree breeding programmes is their rather long time horizon. Tree breeding cycles are often measured in decades rather than years. Compared to breeding programmes of other agricultural crops, some of which can achieve more than one breeding cycle a year, progress in tree breeding is slow and expensive.
Tree breeding programmes have mainly focused on growth, but in another analogy to agricultural crops, growth is not the only requirement for success. A large banana will not sell if it does not taste good. So trees need to grow well but must also produce quality timber. Reasons for neglecting other properties are numerous including negative correlations between growth and some wood quality measures as well as the difficulty to measure wood quality quickly and cost effectively.
The analogy to agricultural crops suggests the strong case for tree breeding for several traits including wood quality. It also highlights the fact that to be successful breeding cycles need to be shortened. The age of selection of superior trees has been reduced in recent years.
Research at the New Zealand School of Forestry developed methods to radically shorten the age of selection for wood quality to an age of one to two years. While it is now possible to make selections of superior trees at a young age, we also need to make sure that the selected trees are quickly propagated. This is to make sure that the seedlings are produced and planted promptly so that they produce an income and to get on with the next breeding cycle. The propagation options for the two purposes may vary depending on the method of tree selection.
Minimising growth strain
Currently the Sustainable Farming Fund is supporting a research programme to minimise growth strain in eucalypts to improve processing. Minimising growth strain in plantation grown eucalypts by breeding will remove one of the most restricting features of these trees for solid wood processing. Around 50 replicates of 200 families which have seed from one tree, a common mother, of Eucalyptus bosistoana and E. argophloia will be screened for growth strain in the next two years.
The newly developed assessment which splits the stem along the pith and measures its distortion is cheap, quick and can be performed when the tree is at age two. However, the test is destructive as it requires the trees to be cut. This has consequences for the propagation of the selected superior trees.


A range of options
There are several strategies for selecting the best trees. It is possible to select the best individuals, the best families or even the best individuals within each family. The strategy which provides the largest gain depends on the heritability − how much of the growth strain variability is under genetic control compared to how much is determined by the environment. Generally, selecting individuals independent of the family works well in most cases. Family selection works best when heritability is low and within-family selection produces the biggest gain when heritability is high.
Another factor to consider is to keep a broad genetic base by including as many families as possible. Selecting best individuals within each family has obvious advantages, while family selection narrows the genetic base the most.
Implications for the eucalyptus breeding programme
What are the implications for the eucalyptus breeding programme? As the assessment of growth-strain is destructive it would seem that families would be an obvious solution as seed from the same mother trees can be collected. This would allow immediate use of reasonable amounts of improved stock and also have the potential to embark quickly on the next breeding cycle by controlled pollination of the mother trees. However, because growth strain is of intermediate heritability, the genetic gain would probably be small and the genetic base of the next generation be drastically reduced. To achieve it all − good genetic gain, a broad genetic base and timely use of improved material − we need to select and propagate individuals which are either independent of families or within families.
We therefore need to retrieve selected individuals following growth strain assessment. It is fortunate that the chosen eucalypts E. bosistoana and E. argophloia coppice, meaning that when they are cut down new shoots grow from the base of the trunk. Coppice growth from the superior individuals can be harvested and grown as cuttings. If, like radiata pine, the species did not coppice, individuals could be kept alive by cutting them above the lowest whorl. The developing branches produce material for propagation. Alternatively, there is the more expensive option of tissue culture. But successful tissue culture has been developed for only a very limited number of tree species.
Vegetative propagation or seeds
Improved material can be used by either of two methods. One is bulk vegetative propagation of selected individuals from cuttings or the other is seed from seed orchards. Clonal propagation has been adopted for several eucalypt species in commercial forestry and can be scaled up quickly. The detail of the procedures for our species needs to be worked out as they are not well researched. This is the approach we are trying first
Crossing superior individuals in a seed orchard can be expected to produce even larger genetic gains, especially when parents originate from very distant populations which have had no opportunity to interbreed. However, there is a considerable lag time until improved material is available as trees need to be old enough to flower and large enough to produce useful quantities of seed. Trees might need to grow for a decade before they start flowering. Nevertheless, the programme also includes seed orchard establishment.
Finally, all this leaves a need for controlled sexual reproduction to begin the next breeding cycle. Sexual reproduction in trees requires them to flower at the same time. Alternatively, male parent pollen needs to be collected, stored and applied to the female parent flowers when they are receptive. Synchronised flowering is not certain considering the distance of the original seed stands of the breeding populations. Grafting improved material to older trees can prompt early flowering. The application of certain hormones might also be possible. The latter is currently being considered at the University of Canterbury.
Logistics
We face a logistic challenge in the current programme because the field trial at Woodville and the propagation facilities in Amberley are in two separate locations.
Live plant material needs to be transported from one site to the other. One option is to cut the coppice and transport the cuttings quickly in cooled containers. As an alternative we have investigated the possibility of lifting the root stools and transporting them to the propagation facilities. This could be an attractive option as the robust root stools are easy to transport as long as they have no coppice and if kept alive would provide a continuous supply of coppice material at the propagation facility.
Root stools from six half-sibling families were lifted over the past growing season. At the beginning of September, October and November a total of 16 plants were lifted and placed into 10-litre buckets.

Bottom: Coppice after five weeks
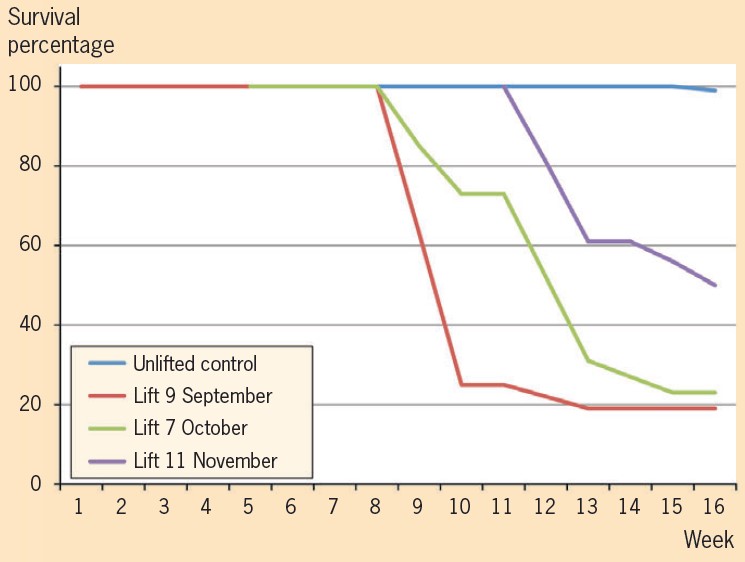
After five weeks 93 per cent were growing coppice shoots. But after the sixth week the shoots started to look unhealthy and plants were starting to die. After three months, only 25 per cent of the lifted plants had survived. However, the plants which survived the first three months are continuing to grow. In comparison the survival rate for the control trees left in the ground is close to 100 per cent.
To ensure that the high loss of plants is not caused by root diseases, plants lifted at the later stage were root pruned and dipped in phosphoric acid for sterilisation, but this treatment did not improve survival. However, a few trees which were put into hydroponics where the medium was much better aerated and they were supplied with ample supply of nutrients, did show an improved survival rate.
The conclusion drawn from this is that damage to the root system by our rough lifting procedure was too severe. Most of the vital fine roots were lost which E. bosistoana could not cope with.

The good point about this is that E. bosistoana appears to be easier to clear from a site. Nevertheless, the E. bosistoana root stools have enough stored energy to produce coppice growth for about a month before running out of energy and succumbing to disease. This might be just enough to harvest cuttings for propagation but without a backup if propagation fails. It might have been that a more gentle lifting procedure or conditioning plants by undercutting to ensure the development of a compact root system, commonly practised in nurseries before transplanting trees, would have produced a better result. Unfortunately there was not enough time to investigate this further.
We have also checked how long it is possible to store living coppice material. It turned out that E. bosistoana coppice is quite resilient. Cut material which was kept for up to a week in a fridge was still vigorous and good propagation success was achievable.
Propagation
The number of coppice shoots per plant is an important factor for propagation. No significant difference in rate of shoot production between lifted and control plants was found. However, shoot production stopped with an average of 27 shoots for each plant lifted after five weeks. This was the time when they started to die.
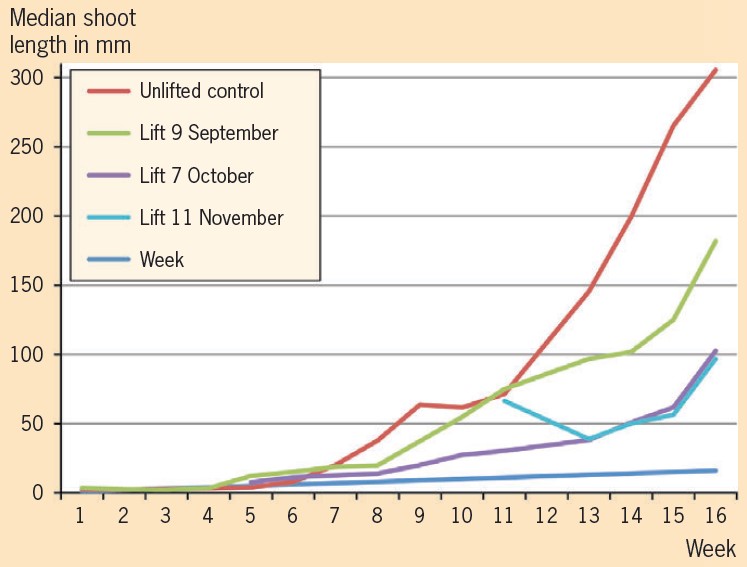
Shoot production continued for plants left in the ground and reached 35 shoots per plant after seven weeks. The number of shoots was also strongly family dependent as well as weakly affected by plant size. Over the long term the shoot growth was more vigorous in the control plants left in the ground than that of the transplanted trees. Coppice shoot growth was not strongly family dependent and independent of plant size.
The cuttings of varying coppice age were set under a range of different conditions. Propagation success was higher with older material. With some treatments propagation success of up to 90 per cent was achieved.
We are now confident that we will be able to propagate superior E. bosistoana trees. Then we will verify the details of the propagation process for E. argophloia, the other species in the current programme.
Paul Schroeder is from Proseed in Amberley and Clemens Altaner from the NZ School of Forestry in Christchurch.